
Another ThreadReader unroll of a recent Twitter thread that I did.

I got several questions from other scientists who all are interested in how to detect science misconduct and how to report. So here is a new thread.
First of all, this is a risky business. Reporting misconduct close to home (e.g. in your own lab or by a close collaborator) might damage not only other people’s careers, but also risk your own job, especially if you are early career.
Second, you need to be able to objectively word your concerns. Yelling “misconduct!” is not going to bring you very far. You have to stick to facts.
“These 2 protein bands look unexpectedly similar” is good.
“These 2 protein bands have been copied and pasted” is subjective.
Third, you have to be patient. If you report a paper/set of papers to a journal or institute, the investigation might take years to complete. Of the 1016 papers, I reported to journals in 2014 and 2015, 54 have been retracted, 181 have been corrected. The rest? … crickets.
Let’s assume that after these 3 disclaimers, you are still interested. There are a couple of extreme scenarios. Maybe you are generally interested in how to spot misconduct and how to respond. Or maybe you suspect misconduct in your lab and are not sure what to do.
If you are generally interested in cases of science misconduct, there are a couple of places you can start:
@Pubpeer is the place where people can comment on a paper – anonymously or signed – positive or negative. Check there regularly for the types of comments people leave.
Also, @PubPeer has a great Chrome plugin that will flag papers (based on DOI) on e.g. Pubmed, so you can see that in your literature searches. It does not seem to work with Google Scholar, unfortunately.

Here is how the Chrome extension will work with Pubmed searches. It will show which papers have comments in @PubPeer .
If you found a paper and you have concerns, you can leave a comment yourself, either under your full name (not recommended when you are just starting) or anonymously (will be moderated, so comments don’t appear immediately).
Make sure to remain objective in your comments. Again, stick to wording such as “unexpected similarities” or “sharp transition between two adjacent bands” instead of “cloned” “fabricated” “manipulated” etc. Assume there is a slight chance that it was an honest mistake
Most of the problems that are found in biomedical papers are potential duplications of photographic images. With my coauthors @ACasadevall1 and @FangFerric, we wrote about those types of problems here (apologies for the weird associated photo):
But there are many other potential problems that you might spot in a paper. You can check for plagiarism by taking part of a sentence (5-8 words works well) between quotes, and searching in Google Scholar. See if you get a single result, or re-use of the sentence.
Definitions will of course give many results, so that is not plagiarism. Here is an example of a definition sentence that will give many results in Scholar: “”Probiotics are live organisms that, when administered in adequate amounts”. That is NOT plagiarism.
But multiple hits in Google Scholar with a sentence such as “”Some of these health problems include bone loss, muscle atrophy, cardiac dysrhythmias” could be cause of concern, especially if there are many other sentences in a paper with multiple hits.
Posting on Pubpeer is one thing, but the conventional – albeit much more slow and ineffective – way of reporting papers with concerns is to write to the Editor of the journal in which the paper was published. Or to the institution in case of multiple papers by the same lab.
Most journals have information on their website with their contact information and their Editor in Chief. It is most effective to write to more than one email address (pick a couple of senior editors as well) so that there is more chance that a journal will actually respond.
I always write per email, not paper letters, as to leave a record. Unfortunately, some journals make it very hard to find their contact information. You might have to cyberstalk the editors and search Pubmed publications or faculty pages for their email addresses.
If there are multiple problematic papers, you can also report to the institution / university. There might even be multiple institutions, if a person moves from lab to lab. Search for “Research Compliance” or “Research Integrity” and the university’s name. It might be hidden.
How about if you suspect misconduct close to yourself (e.g. by a co-worker in your lab)? If you trust the PI, you could first raise it with them. If not, you could report it to the Research Integrity office of your university. Write them an anonymous (paper) letter or email.
Unfortunately, it is very risky to write under your own name. Research Integrity officers might promise you anonymity, but might reveal your name to the defendant in a later stage of the investigation. This has happened to me and it sucks.
That is just some general advice that I have about how to spot and report cases of misconduct. Happy to talk more about this in a different thread.
My upcoming year of pro bono research integrity work
Hi all,
I have decided to take (at least) a year off from paid work to focus on my research integrity work. Since 2013, I have worked on finding plagiarism and image duplication in scientific papers. Every free minute I searched for papers, made reports highlighting the potential problems, and wrote to journals and institutions about these concerns.
Together with Arturo Casadevall, Ferric Feng and other co-authors, we published three papers about our work on the frequency of image duplication in biomedical papers. You can find them here:
- The Prevalence of Inappropriate Image Duplication in Biomedical Research Publications – Elisabeth M. Bik, Arturo Casadevall, Ferric C. Fang – mBio. 2016 Jun 7;7(3). pii: e00809-16. doi: 10.1128/mBio.00809-16
- Testing Hypotheses on Risk Factors for Scientific Misconduct via Matched-Control Analysis of Papers Containing Problematic Image Duplications – Daniele Fanelli, Rodrigo Costas, Ferric C. Fang, Arturo Casadevall, Elisabeth M. Bik – Sci Eng Ethics. 2018 Feb 19. doi: 10.1007/s11948-018-0023-7
- Analysis and Correction of Inappropriate Image Duplication: the Molecular and Cellular Biology Experience – Elisabeth M. Bik, Ferric C. Fang, Amy L. Kullas, Roger J. Davis, Arturo Casadevall – Mol Cell Biol. 2018 Sep 28;38(20). pii: e00309-18. doi: 10.1128/MCB.00309-18
As of today, I have reported 85 papers and theses for extensive plagiarism, and over 1200 papers for potential image duplication. But I have many more that I still need to report, and I am getting more and more requests for help, and the work – all done in my spare time – was piling up.
So after working for 2.5 years in microbiome startup companies, I have decided to take at least a year off – maybe longer – so that I will have much more time to work on this project. I will still closely follow the microbiome field, of course, as well.
Thank you all, for your support!
Elisabeth Bik
Here is the unroll of a Twitter thread on this topic, as done by ThreadReader.

I am taking a year off from paid work to focus more on my science misconduct volunteer work. Science needs more help to detect image duplication, plagiarism, fabricated results, and predatory publishers.
Most of the work detecting these problems in science papers is done by volunteers like me. It takes perseverance and patience. Many journals, authors, and academic institutions will not take action.
Even if they respond, It might take years before papers with serious flaws are corrected. All that time, those papers are not flagged by the journals, and others researchers might cite them or base their research on them.
As of now, we can only flag papers on @PubPeer and install their plugin so you can see which papers have a comment, e.g. when doing literature searches.
And I will still write to journals or institutions about all papers with concerns that I found so far. Even if it takes hours to find their contact info.
I still have 100s of papers that I need to officially report and 100s of reported ones to follow up on. The only way I felt I can catch up on that is to quit my paid job. Which is scary.
It would be nice if journals, institutions, funding agencies, and countries would care more about the quality of their research. If they had more guts to respond to concerns raised by readers – and take action.
The work that volunteers like us do is not very rewarding obviously. No one likes criticism. It can also be dangerous. Authors might start personal attacks on us and sue us for libel.
I am also very aware of the collateral damage, e.g. of coauthors who did not commit misconduct, others workers in those labs, and the effects on family members.
With that in mind, it is important to focus on facts, on the potential problems in the papers and how to address those. The focus should be on the papers, not the authors.
I might make an exception in cases of authors using false affiliations and fake coauthors.
The more you dig into these cases the more other weird stuff you find. I can probably do this full-time for the rest of my life. Maybe I will.
A review of the recent Viome preprint
An open peer review of a preprint paper by Hatch A et al. from Viome, posted on OSF Preprints, January 2019.
Full disclosure: I worked for Viome’s competitor uBiome from October 2016 – December 2018. I am currently an independent consultant.
Andrew Hatch, James Horne, Ryan Toma, Brittany L. Twibell, Kalie M. Somerville, Benjamin Pelle, Kinga P. Canfield, Guruduth Banavar, Ally Perlina, Helen Messier, Niels Klitgord and Momchilo Vuyisich*
Viome, Inc, Los Alamos, NM 87544, United States.
Background information
In a recent paper authored by Stanford researchers, biotech startups were criticized for not sharing their discoveries through peer-reviewed research studies. So in that light, it is great to see a biotech company such as Viome publish a study about their microbiome consumer product. Viome’s leaders have been very vocal about the superiority of their product – which is based on RNA transcription – over that of other microbiome consumer tests, which are based on DNA amplification and sequencing. But until this preprint came out, no research on the Viome product had been published. So I was excited to hear about this preprint!
It is important to first point out that this paper is not a peer-reviewed paper. It is a preprint, which means it is written as an academic paper, but it has not been peer-reviewed by other scientists. It is a first step, however, to share the work that Viome did to build a metatranscriptomics platform, and show some of their first results. I hope my comments will be useful in the process of getting this study published in a peer-reviewed journal.
The paper describes Viomega, Viome’s automated stool metatranscriptomics method that involves RNA extraction from stool samples, sequencing, and bioinformatics analysis. Let’s go over each of the sections.
 Introduction
Introduction
The introduction of the paper is mostly stating how metatranscriptomics is superior to other techniques. It has a somewhat oversimplified table showing other methods (bad!) to their own method (good!).
For example, under “Method Biases”, the 16S Gene Sequencing column states “Heavily influenced by amplification method, but also sequencing quality, sample lysis, and bioinformatics“, where “heavily influenced” sounds a bit denigrating, while sequencing quality and bioinformatics are conveniently left out in the Metatranscriptomics column.
The “identifies all living organisms” appears oversimplified as well. First, is a phage or virus alive? Then, how about a bacterium that is in a viable-non-culturable or spore-state? It is alive, but it is probably not transcribing much RNA – can metatranscriptomics detect those?
The statement that Metatranscriptomics “allows assessment of pathway activities that can lead to personalized health insights and recommendations with molecular-level precision” is overly subjective and is not proven in this paper. That last part of the sentence sounds like a Viome commercial, not like something that belongs in a scientific paper.
The introduction text states that 16S gene sequencing misses archaea or eukaryotes, but fails to acknowledge that most of these can be identified with broad-range primers as well.
In short, the Introduction shows a too black- and white comparison between different microbial community methods that is not very objective.
Methods
Not unexpectedly from a biotech company, the paper does not provide a lot of technical details on sample extraction, library preparation, or bioinformatics analysis. In order to pass peer review, the authors will likely need to provide more details on their methods – so that others can easily replicated them. The statements about participant consent and IRB approval were also very short; most journals and peer reviewers would like to see something more than “all study procedures were approved by an IRB” from a non-academic institution.
Results: Validation and reproducibility
The first part of the Results shows that the Viomega method can detect a range of different microorganisms with relative equal efficiency and precision. Using mock communities, the results are accurate, and reported to contain no false positives or negatives or sample-to-sample crosstalk. However, data given here were very sparse.
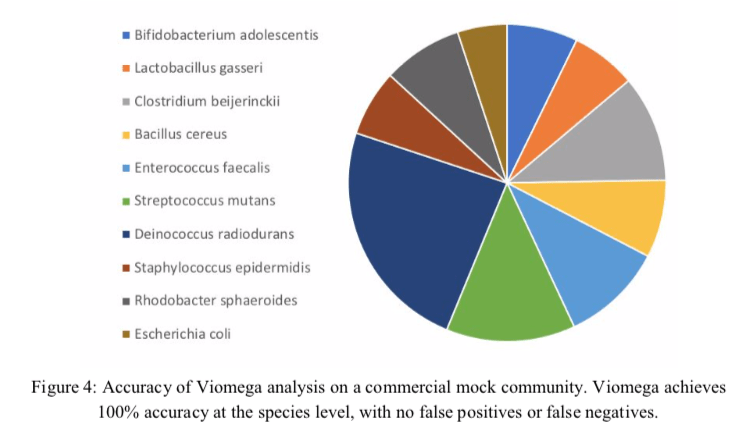
Figure 4 is a colorful representation of the mock community sequencing experiment, but is lacking percentages and other details, such as number of reads. Where there really no false positive or false negative reads? Not a single one? There is also no word on how the negative controls did perform over their tests of 10,000 samples. It would have been really valuable to have compared this mock community using all three methods compared in Table 1; 16S, metagenomics, and metatranscriptomics.
This section of the Results also shows that the Viome test is reproducible. Three different experiments were performed in which small numbers of participants (3 to 7 persons) collected stool samples testing the following:
- stool collection from different part of their stool sample (Figure 5)
- waiting 0, 3, or 10 min before adding stool to the kit tube (Figure 6)
- collecting stool on 3 different days over a period of two week (Figure 7)
In all three experiments, samples from the same stool specimen or individual were very similar to each other, showing that the Viome test results are reproducible (the dark squares along the diagonal in Figures 5, 6, 7).
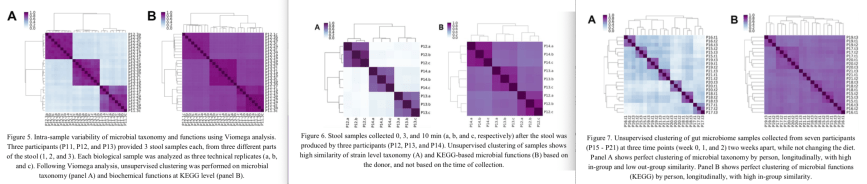
Is Gene Function Composition better than simple Taxonomy?
Figures 5/6/7 show the test results in two different ways. The A panels in each figure are based on which active microbes are present in the samples, so based on the taxonomy of the RNA reads. The B panels are the results based on gene function composition (which genes are being expressed in that sample).
It was interesting to see that in all cases the microbial composition (taxonomy; A panels) was better able to tell individuals apart than the gene expression (B panels).
Detailed explanation: If you look at the A panels, the comparison of each person to other samples from the same person shows they are very similar (purple) while each individual is very different (light blue) from other persons. In the B panels, the individuals do not differ that much from each other; all individual-to-individual comparisons are darker shades of purple that are more difficult to tell apart.
That is a bit ironical, because the preprint states at the beginning (Abstract and Table 1) that functional gene analysis, not microbial composition based on taxonomy, is needed for personalized health insights. Instead, the paper appears to show that each person has their own, personal gut microbiome, while the functional capacities appear to be pretty similar between individuals.
The paper does not provide an answer how these small person-to-person variations in microbial gene expression will lead to the “goal to develop personalized nutrition algorithms” as stated in the Abstract.
Transcriptomics taxonomy prevalence data from 10,000 samples
Metatranscriptomics is superior to other techniques, stated the paper in the Introduction and Table 1, but the paper does not do anything to prove that. There was no comparison of 16S sequencing vs metatranscriptomics on e.g., the mock community shown in Figure 4. Such a comparison could add a lot of value to the paper, and could support the bold statements made in the Introduction.
Also, the amount of viruses, archaea, and eukaryotes in the sample set was not very high, suggesting that 16S sequencing does not miss as much diversity as the authors claim in the introduction. For example, strain level analysis (Table S1) shows crAssphage as the most prevalent virus, but it is ranked #144 of all taxa (26%). Similarly, Methanobrevibacter clocks at #269 (14%), while Entamoeba is the most prevalent eukaryote at #336 (10% of samples).
Many of the viruses/phages found in the stool samples appear to be plant associated (Suppl tables). For example, Phaseolus vulgaris endornavirus (beans), Pepper mild mottle virus, Cannabis cryptic virus, and shallot latent virus are among the most prevalent viruses. Could the authors comment on this? Could these be transient microbiome components that were part of a food item, or are they present in the same person over longer periods of time? It would be nice to see some data on that.
Several taxa appear to behave very differently on strain/species/genus level (Tables S1-S3). Examples where genus-level prevalence is much lower than expected based on strain prevalence are given below. In other examples, taxa with high prevalence at genus level seem to disappear at species or strain level; also given below.
Unfortunately, relative abundance data appears to be missing. How abundant are viruses, archaea and eukaryotes ? Which percentage of the transcriptomics reads were assigned to each of these groups? These numbers, now missing, would tell us which groups 16S sequencing would miss and would allow for a better comparison of different microbial community analysis tools.
Can Viome’s microbiome analysis connect the diet and the gut metatranscriptome?
The paper contained very few functional results on the data from 10,000 Viome samples. Viome has repeatedly claimed that transcriptomics are much more informative than just lists of microbial taxa, and the abstract promises “several small clinical studies to demonstrate the connections between diet and the gut metatranscriptome.”
Therefore, it was disappointing to see that the paper was mainly limited to taxonomic assignments. This dataset, which is one of the largest of its kind, sounded very promising, and I had hoped to see many more functional analyses on these samples.
Table 4 is the only part of the study that analyzes the functional capacity of the data from 10,000 samples. It lists the top 12 KEGG functions (although the legend says “top 10”; see below). Unfortunately, this table is not very informative and might contain some errors (see below). Most importantly, it is just a list of genes without any discussion on their function.
In addition, if the prevalence of these genes is over 99.9% (meaning that almost every person’s microbiome contains those genes), how can one use that for the correlation of microbial taxa or genes with lifestyle or diet? Based on the top 10 or top 100 (Table S4) KEGG functions, all subjects’ microbiomes appear to have the exact same genes. How is this functional data superior to the big inter-individual differences that are found by the more conventional 16S types of microbiome analyses? The paper would be much stronger if this important point would be discussed.
Summary of findings
In summary, the strengths of this paper are the experiments showing that the Viomega technique is reproducible. That is a great paper by itself, but the current title and abstract are promising much more than the paper currently delivers. With a transcriptomics dataset of 10,000 stool samples, and repeated claims that transcriptomics will give much more functional insight than just a list of taxa, it is disappointing to see that the results are limited to microbial taxon prevalence and not any functional analysis.
Based on this paper, Viome’s claims that their test can connect the microbiome transcriptome to a subject’s diet – let alone give dietary advice – appear to be very far-fetched.
Specific / detailed comments
- Introduction: Reference 1, Nasca 2018, does not appear to be about the microbiome but about Acetyl-L-carnitine deficiency in people with depression
- “16S gene” should be “16S rRNA gene”, at least the first mention in the manuscript
- Not all 16S sequencing uses a short piece, in several cases one can identify to species level, depending on length of gene fragment used
- “In addition, it misses some bacteria, most archaea, and all eukaryotic organisms and viruses [19].” Actually, reference 19 shows that great archaeal diversity can be captured using archeaon specific 16S primers (which is still 16S sequencing) – as long as you use different primers. Also this reference does not support the statement that 16S sequencing misses eukaryotic organisms and viruses.
- Table 1 is not very objective (see above). The last 2 rows appear to say the same thing; statement in the right/bottom cell is too strong. Top right cell: only if metabolically active.
- Results: the first section (“Validation…”) might be better moved to the Methods.
- Figure 1. It is not quite clear what the input here was. The RNA yield is given, but how were the input amounts relative to each other? Were equal number of cells added, or equal amount of turbidity? Were samples normalized for e.g. genome size?
- “Ambient sample transportation”: The statement “There has been unfounded criticism of metatranscriptomics ” does not appear to be very scientific, and it lacks a reference.
- Figure 2: It is not clear which of the peaks shows the RNA. The peaks in B are much lower; it would be better to show both panels at the same Y-axis scale. Also, if these are two “representative” samples, did they contain the same original sample?
- Page 6: “Viomega eliminates STSC altogether” is not true – while it is low, it is not 0, as shown in Figure 3, so this needs to be reworded.
- Figure 3: “Cross talk” is a very technical term – the figure legend should define how this is measured (text in Results was more helpful).
- Page 7: “more than 1,000 fold lower ” – this data is not shown – either provide a reference or do the experiment.
- Page 7, “Background contamination”. “Since any metagenomic or metatranscriptomic analysis identifies all taxa in a sample” is only true for live, transcribing organisms.
- This section also states that background contamination would be picked up by the PPC sample (positive control) but is much more likely to be detected by the negative control sample. However, the study does not show how many reads were detected in the NC samples. Could the authors add that data? The Salter paper clearly shows that background taxa are rarely detected in samples with high amount of “real” taxa – they only show up when there is very low amount of real nucleic acid to detect.
- What is Table 2 showing? Number of contaminating reads in PPC samples? More detail is needed here. How can these 1.4 reads per sample be distinguished from the “Sample to Sample Cross Talk” reads? Both Figure 3 as well as Table 2 appear to be studying the same thing, that is the number of reads that were assigned to different taxa than included in the PPC mixture. Could the authors clarify the difference?
- “Findings from the Viomega taxonomic classification”. The main text says that Tables S1-S3 include the top 100 taxa, but this appears incorrect. E.g., S1 lists 2723 taxa (strain level), so there are many more taxa than 100.
- Table 4
- The table lists 12 KEGGS, so it is not a top 10 as mentioned in the text
- The top 10 KEGGS (table 4) and the top listings in Table S4 do not match. K00540 (#5 in Table S4) K03296 (#7) and K06950 (#10) are not present in Table 4. In contrast, K03737 (#3 in table 4), K00975 (#8), K00705 (#9) are not present in the top 10 in Table S4.
- Tables 4/S4: a short description of what these functions entail would be welcome.
- KEGG K00936 appears to be a two-component system, sensor histidine kinase PdtaS [EC:2.7.13.3] – not E2.7.3.-[EC:2.7.3.-] as denoted.
- Page 11. Subject 11’s stool appears to show some clustering per stool section (1/2/3). “Biological replicates were indistinguishable from the technical replicates.” might therefore be incorrect; some statistical analysis is needed (.e.g Lin’s correlation) to see which factor (technical vs biological replicate vs subject) is the strongest.
- Page 11. “While there have been claims of large intra-sample variability” – this refers to a paper (#34) that tested samples from different persons and different anatomical sites which – not surprisingly – were found to vary. However, in the current study, the authors tests biological and technical replicates, something that was not tested in Reference 34.
- Tables S1-S3: Several taxa appear to behave very differently on strain/species/genus level.
Examples where genus-level prevalence is much lower than expected based on strain prevalence:
Eggerthella
Strain: Eggerthella lenta 1_1_60AFAA, at 97.08% prevalence
Species: Eggerthella lenta at 91.76%
Genus: Eggerthella at 61.12%
Veillonella
Strain: Veillonella dispar A at 92.34
Species: Veillonella dispar at 88.99%
Genus: Veillonella at 78.62%
Entamoeba:
Strain: Entamoeba nuttalli P19 at 9.85% / Entamoeba dispar SAW760 0.74%
Species: Entamoeba nuttalli: 9.87% / Entamoeba dispar 0.74%
Genus: Entamoeba: 0.79%
In other examples, taxa with high prevalence at genus level seem to disappear at species or strain level:
Lactococcus
Strain: Lactococcus piscium MKFS47 at 0.99% / Lactococcus raffinolactis NBRC 100932 at 0.23%
Species: Lactococcus piscium at 0.98%
Genus: Lactococcus at 96.88%
Escherichia
Strain level: Escherichia coli isolate 15 at 37.8%; Escherichia coli strain LS5218 at 9.5%, Escherichia coli M17 at 7.7%.
Species level: Escherichia coli at 27.40%
Genus level: Escherichia: 85.92% prevalence
Saccharomyces
Strain: Saccharomyces sp. ‘boulardii’ strain unique28 is present at 1.82% of samples, as the most prevalent Saccharomyces strain. Also Saccharomyces cerevisiae S288C at 0.80%.
Species level: S. cerevisiae is present in 3.9% of samples.
Genus level: Saccharomyces it is present in 25.44%.
Streptococcus
Strain: Streptococcus sp. 263_SSPC 5.56% / Streptococcus mutans U138 at 1.62%
Species: Streptococcus thermophilus 28.15% / Streptococcus mutans 9.92% / Streptococcus parasanguinis 7.54%
Genus: Streptococcus 90.47%
Salmonella
Strain: Salmonella enterica subsp. enterica strain SE696A 2.30%; Salmonella enterica subsp. enterica strain ADRDL-LA-5-2013 2.28% / Salmonella enterica subsp. enterica serovar Typhimurium strain 1.52%
Species: Salmonella enterica 11.71%
Genus: Salmonella 15.61%
April 25, 2019
Good morning everyone! (and apologies for the delay, a technical issue regarding the system time difference between Israel and US Pacific time :)).
Due to yesterday extremely rich digest, today’s post will be short- but diverse! Spreading from gut microbiome in infants and beetles, to the effect of microbial communities on sweet wine quality. As a bonus for the upcoming weekend, at the end of the post you’ll find a link to an article listing 10 recommended biotech podcasts.
Happy holidays for all!
General Science and Microbiology
Failure to vaccinate and vaccine failure– Editorial- Nature Microbiology
The problem with unadjusted multiple and sequential statistical testing– Casper Albers- Nature Communications
Human microbiome
**Review – What did we learn from multiple omics studies in asthma?– Olga Ivanova- Allergy
Fecal microbiota transplant for Crohn disease: A study evaluating safety, efficacy, and microbiome profile– Liat Gutin- ueg
Specific class of intrapartum antibiotics relates to maturation of the infant gut microbiota: a prospective cohort study– Modupe O. Coker- BJOG
The Gut Microbiome on a Periodized Low-Protein Diet Is Associated With Improved Metabolic Health– Zhencheng Li- Frontiers in Microbiology
Plant microbiome
The plant host induces antibiotic production to select the most beneficial colonizers– Ariel Ogran- AEM
Microbial network and soil properties are changed in bacterial wilt susceptible soil– Gaufo Qi- AEM
Water Microbiomes
Fates of antibiotic resistance genes in a distributed swine wastewater treatment plant– Qing-Bin Yuan- Water Environment Research
** Metabolic potential of uncultured bacteria and archaea associated with petroleum seepage in deep-sea sediments– Xiyang Dong- Nature Communications
Animal microbiome
Gut anatomical properties and microbial functional assembly promote lignocellulose deconstruction and colony subsistence of a wood-feeding beetle– Ceja Navarro- Nature Microbiology
Gut microbiome-derived phenyl sulfate contributes to albuminuria in diabetic kidney disease– Koichi kikuchi- Nature Communications
Honeybee-specific lactic acid bacterial supplements have no effect on American foulbrood infected honeybee colonies– Jorg G. Stephan- AEM
Food microbiome
Influence of microbial communities on the chemical and sensory features of Falanghina sweet passito wines– F De Fillips- Europe PMC
Microbiome in the News
The Asia Pacific human microbiome market is expected to reach US$ 207.81 Mn in 2025 -Cision
**UPDATED- Critical Reading required– Antibiotics: beneficial side effects are starting to come to light– Claire Bourke- The Conversation. I’m bringing about this post in order to encourage a discussion regarding the issues and claims that were written in this non-scientific article. You’re welcome to share your thoughts about it at the comments section below.
Podcasts
Top 10 Biotech Podcasts to Listen to this Spring– Clara Rodriguez Fernandez- Labiotech.eu
**Personal recommendation- This Week in Microbiolgy (TwiM)
April 22, 2019
(Posting for our new team member, Zehra Tüzün Güvener.)
Good Morning! Highlights from Monday’s digest include a widely distributed efflux pump in saprophytic soil bacteria, genomic analysis of plant-growth-promoting rhizobacteria, identification of novel mammalian viruses from bats, rectal swabs to study gut microbiomes and TrueBiome’s solution to reduce variability in animal experiments.
General Microbiology
Efflux pumps in Chromobacterium species and their involvement in antibiotic tolerance and survival in a co-culture competition model – Benomar et al. – BioRxiv
Muropeptides Stimulate Growth Resumption from Stationary Phase in Escherichia coli – Joers et al. – BioRxiv
Human oral microbiome
Current Understanding of the Gut Microflora in Subjects with Nutrition-Associated Metabolic Disorder Such as Obesity and/or Diabetes: Is There Any Relevance with Oral Microflora? – Yumoto et al. – Current Oral Health Reports
Human skin microbiome
Scalp bacterial shift in Alopecia areata – Pinto et al. – PLOS ONE
Human gut microbiome
Factors influencing the gut microbiome in children: from infancy to childhood – Kumbhare et al. – Journal of Biosciences
Animal microbiomes
Defining Escherichia coli as a health-promoting microbe against intestinal Pseudomonas aeruginosa – Christofi et al. – BioRxiv
Glyphosate, but not its metabolite AMPA, alters the honeybee gut microbiota – Blot et al. – PLOS ONE
Plant, root and soil microbiome
**MiniReview: Gene mobility in microbiomes of the mycosphere and mycorrhizosphere –role of plasmids and bacteriophages – Pratama et al. – FEMS Microbial Ecology
**Review: Response of microbial communities to biochar-amended soils: a critical review – Palansooriya et al. – Biochar
The effects of soil phosphorous content on microbiota are driven by the plant phosphate starvation response – Finkel et al. – – BioRxiv
Comparative genomic analysis of Bacillus paralicheniformis MDJK30 with its closely related species reveals an evolutionary relationship between B. paralicheniformis and B. licheniformis – Du et al. – BMC Genomics
Are drivers of root-associated fungal community structure context specific? – Alzarhani et al. – The ISME Journal
Water and extremophile microbiome
Impact of NaCl, nitrate and temperature on the microbial community of a methanol-fed, denitrifying marine biofilm – Villemur et al. – BioRxiv
Metabolic potential of uncultured bacteria and archaea associated with petroleum seepage in deep-sea sediments – Dong et al. – Nature Communications
Probiotics / prebiotics
Screening of single or combined administration of 9 probiotics to reduce ammonia emissions from laying hens – Mi et al. – Poultry Science
Phages and viruses
A viral metagenomic survey identifies known and novel mammalian viruses in bats from Saudi Arabia – Mishra et al. – PLOS ONE
Techniques
Metaproteomics of Freshwater Microbial Communities – Russo et al. – Mass Spectometry of Proteins (Methods in Molecular Biology)
Functional annotation of orthologs in metagenomes: a case study of genes for the transformation of oceanic dimethylsulfoniopropionate – Gonzales et al. – The ISME Journal
Usability of rectal swabs for microbiome sampling in a cohort study of hematological and oncological patients – Biehl et al. – PLOS ONE
Microbes in the news
** Report – North America Human Microbiome Market to 2025 – Regional Analysis and Forecasts by Product, Disease, Application, and Country – Research and Markets
Microbes on the market
** Newsletter – Addressing experimental variability: Taconic launches TruBiome – Outsourcing-Pharma.com
April 16, 2019
Good morning everyone! Today’s digest will deal a lot of disturbance in various ecosystems- starvation of the oral microbiota, forest fires, elevated CO2 levels and rising temperatures and more. Additionally, interesting pieces about science communication and education are recommended (bottom of the post).
Animal microbiome
Effects of diet on gut microbiota of soil collembolans– Qian Xiang- Science of the Total Environment
Diel population and functional synchrony of microbial communities on coral reefs– Winda Wegley-Kelly- Nature Communications
Human microbiome
Klebsiella and Providencia emerge as lone survivors following long-term starvation of oral microbiota– Jonathon L. Baker- PNAS
Horizontal gene transfer in human-associated microorganisms inferred by phylogenetic reconstruction and reconciliation– Hyeonsoo Jeong- Scientific Reports
Dysbiosis patterns during re-induction/salvage versus induction chemotherapy for acute leukemia– Armin Rashidi- Scientific Reports
The personalized temporal dynamics of microbiome in the airways of cystic fibrosis patients– Giovanni Baci- bioRxiv
Plant, root and soil microbiome
Maize synthesized benzoxazinoids affect the host associated microbiome- Enoch Narh Kodjordjie- Microbiome
Utilization of carbon sources in the rice rhizosphere and non-rhizosphere soils with different long-term fertilization management– Tang H- J of Basic Microbiology
Effect of forest fire prevention treatments on bacterial communities associated with productive Boletus edulis sites- Olaya Mediavilla- Microbial Biotechnology
Effects of elevated temperature and elevated CO2 on soil nitrification and ammonia-oxidizing microbial communities in field-grown crop– Linh T.T Nguyen- Science of the Total Environment
Phyllosphere epiphytic and endophytic fungal community and network structures differ in a tropical mangrove ecosystem– Hui Yao- Microbiome
Changes in fungal communities across a forest disturbance gradient– Lingling Shi- AEM
Water microbiome
Wetland sediments host diverse microbial taxa capable of cycling alchohols- Paula Dalcin Martins- AEM
Review- The microbiomes of deep-sea hydrothermal vents: distributed globally, shaped locally– Gregory J. Dick- N. Reviews Microbiology
Bioinformatics
DeeplyEssential: A Deep Neural Network for Predicting Essential Genes in Microbes- Md Abid Hassan- bioRxiv
Microbiome news and science communication
Biotech company launches animal models with custom microbiome profiles
Microbial biotechnology for water treatment- A collection of high-quality websites (from Microbial Biotechnology)
Thematic series on scholarly communications in the digital age
8 April, 2019
General Microbiome
Coevolutionary dynamics shape the structure of bacteria-phage infection networks. Fortuna MA.Evolution.
High proportions of bacteria are culturable across major biomes. Martiny AC. ISME Journal
The complexity of an optimal vaginal biome. Eschenbach DA. BJOG
The lung microbiome, vitamin D, and the tuberculous granuloma: A balance triangle. Balcells ME. Microbial Pathogenesis
Gut Microbiome
The parasitic worm product ES-62 normalises the gut microbiota bone marrow axis in inflammatory arthritis. Doonan J. Nature Communications
Culture-enriched human gut microbiomes reveal core and accessory resistance genes. Raymond F.Microbiome.
Plant, Soil, Water, and Air Microbiome
Temperature and soil moisture control microbial community composition in an arctic-alpine ecosystem along elevational and micro-topographic gradients.Frindte K. ISME Journal
Root traits and belowground herbivores relate to plant-soil feedback variation among congeners.Wilschut RA. Nature Communications
A Polyphasic Approach for Assessing Eco-System Connectivity Demonstrates that Perturbation Remodels Network Architecture in Soil Microcosms.Stamou GP. Microbial Ecology
Effect of stock density on the microbial community in biofloc water and Pacific white shrimp (Litopenaeus vannamei) gut microbiota.Deng Y. Applied Microbiology Biotechnology
Rapid and stable microbial community assembly in the headwaters of third-order stream. Teachey ME. Applied Environmental Microbiology
Comparative Metagenomics Reveals Enhanced Nutrient Cycling Potential After Two Years of Biochar Amendment in a Tropical Oxisol. Yu J. Applied Environmental Microbiology
Animal Experiments and Microbiome
Effect of a low protein diet on chicken ceca microbiome and productive performances. De Cesare A. Poultry Science
Impact of a probiotic-based cleaning product on the microbiological profile of broiler litters and chicken caeca microbiota.De Cesare P. Poultry Science
Metagenomic Analysis of the Whole Gut Microbiota in Brazilian Termitidae Termites Cornitermes cumulans, Cyrilliotermes strictinasus, Syntermes dirus, Nasutitermes jaraguae, Nasutitermes aquilinus, Grigiotermes bequaerti, and Orthognathotermes mirim. Grieco MB. Current Microbiology.
The hemolymph microbiome of insects. Blow F. Journal Of Insect Physiology
Musa basjoo regulates the gut microbiota in mice by rebalancing the abundance of probiotic and pathogen. Wei T. Microbial Pathogenesis
Lytou AE. International Journal of Food Microbiology.
April 05, 2019
Good morning everyone! In today’s microbiome digest, we have multiple research and review articles focusing on the role of microbiome in Prostate Cancer, Breast Cancer, Gastric Cancer, Allergy, Gut-Brain axis, Schizophrenia, Paediatric Crohn’s Disease, Nonalcoholic Fatty Liver Disease, Alveolar Osteitis, Renal Failure, Cholecystokinin Expression, Lipid Metabolism Disorder, Regulation of Blood Pressure and an independent section on Fecal Microbiota Transplantation. Additionally, two very interesting metagenomic studies have discovered specific microbial signature as well as link with choline degradation in the colorectal cancer patients. Enjoy reading.
General microbiome
Reivew: The Human Microbiota and Prostate Cancer: Friend or Foe? – Francesco Massari – Cancers
Microbiome—Microbial Metabolome—Cancer Cell Interactions in Breast Cancer—Familiar, but Unexplored – Edit Miko – Cells
Review: Fighting AMR in the Healthcare Environment: Microbiome-Based Sanitation Approaches and Monitoring Tools – Maria D’Accolti – Molecular Sciences
Gut microbiome
Gut microbial dysbiosis is associated with allergen-specific IgE responses in young children with airway allergies – Chih-Yung Chiu – World Allergy Organization Journal
Review: The gut virome: the ‘missing link’ between gut bacteria and host immunity? – Indrani Mukhopadhya – Therapeutic Advances in Gastroenterology
Regulatory T Cells and Plasmacytoid Dendritic Cells Within the Tumor Microenvironment in Gastric Cancer Are Correlated With Gastric Microbiota Dysbiosis: A Preliminary Study – Zongxin Ling – Frontiers in Immunology
Review: Impact of Different Types of Diet on Gut Microbiota Profiles and Cancer Prevention and Treatment – Rainer J. Klement – Medicina
Review: Glutamatergic Signaling Along The Microbiota-Gut-Brain Axis – Andreina Baj – Molecular Sciences
Investigation of the Gut Microbiome in Patients with Schizophrenia and Clozapine-Induced Weight Gain: Protocol and Clinical Characteristics of First Patient Cohorts –Gorbovskaya I. – Neuropsychobiology
Review: Sex Differences in Gut Microbiota – Yong Sung Kim – The World Journal of MEN’s HEALTH
The Microbiome in Paediatric Crohn’s Disease—A Longitudinal, Prospective, Single-Centre Study – Shivani Kansal – Journal of Crohn’s and Colitis
A gut microbiome signature for cirrhosis due to nonalcoholic fatty liver disease – Cyrielle Caussy – Nature Communications
Metagenomic analysis of colorectal cancer datasets identifies cross-cohort microbial diagnostic signatures and a link with choline degradation – Andrew Maltez Thomas – Nature Medicine
Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer – Jakob Wirbel – Nature Medicine
Ceftriaxone and cefotaxime have similar effects on the intestinal microbiota in human volunteers treated by standard doses regimens – Charles Burdet – Antimicrobial Agents and Chemotherapy
Human respiratory microbiome
Mycobiome Sequencing and Analysis Applied to Fungal Community Profiling of the Lower Respiratory Tract During Fungal Pathogenesis – Lisa R. McTaggart – Frontiers in Microbiology
Reproductive tract microbiome
Identification and evaluation of the microbiome in the female and male reproductive tracts – Rivka Koedooder – Human reproduction update
Skin microbiome
Mast cell recruitment is modulated by hairless skin microbiome – Chia-Chi Wu – The Journal of Allergy and Clinical Immunology
Oral microbiome
A metagenomic study of patients with alveolar osteitis after tooth extraction. A preliminary case-control study – Laura Aguilar-Duran – Clinical Oral Investigations
Animal microbiome / Animal experiments
Interactions Between Commensal Bacteria and Enteric Neurons, via FPR1 Induction of ROS, Increase Gastrointestinal Motility in Mice – Bindu Chandrasekharan – Gastroenterology
Danshen can interact with intestinal bacteria from normal and chronic renal failure rats – Hongdie Cai – Biomedicine and Pharmacotherapy
Review: A place for taxonomic profiling in the study of the coral prokaryotic microbiome – Alejandra Hernandez-Agreda – FEMS Microbiology Letters
Fructose malabsorption induces cholecystokinin expression in the ileum and cecum by changing microbiota composition and metabolism – Xufei Zhang – THE FASEB JOURNAL
Spinach consumption ameliorates the gut microbiota and dislipaemia in rats with diet-induced non-alcoholic fatty liver disease (NAFLD) – Elvira-Torales LI – Food and Functions
Microbial Colonization in Adulthood Shapes the Intestinal Macrophage Compartment – Franziska Schmidt – Journal of Chron’s and Colitis
Feeding modes shape the acquisition and structure of the initial gut microbiota in newborn lambs – Yanliang Bi – Environmental Microbiology
Effects of dietary lipid sources on the intestinal microbiome and health of golden pompano (Trachinotus ovatus) – Cuihong You – Fish & Shellfish Immunology
Gut microbiota characterization and lipid metabolism disorder found in PCB77-treated female mice – Chi Y – Toxicology
Changes of gut microbiota between different weight reduction programs – Belle Yanyu Lin – Surgery for Obesity and Related Diseases
Microbiome-metabolomic analysis of the impact of Zizyphus jujuba cv. Muzao polysaccharides consumption on colorectal cancer mice fecal microbiota and metabolites – Xiaolong Ji – International Journal of Biological Macromolecules
Fecal Microbiota Transplantation
Critical Role of the Interaction Gut Microbiota – Sympathetic Nervous System in the Regulation of Blood Pressure – Marta Toral – Frontiers in Physiology
Faecal microbiota transplantation for the decolonisation of antibiotic-resistant bacteria in the gut: a systematic review and meta-analysis – Vera Tavoukjian – The Journal of Hospital Infection
Prebiotics/Probiotics
Effect of a multistrain probiotic (Lactoflorene®Plus) on inflammatory parameters and microbiota composition in subjects with stress-related symptoms – Sara Soldi – Neurobiology of Stress
Probiotics, prebiotics, and low FODMAP diet for irritable bowel syndrome – What is the current evidence? – Soo Liang Ooi – Complementary Therapies in Medicine
Water and extremophile microbiome
Microbial Community of Saline, Alkaline Lakes in the Nebraska Sandhills Based on 16S rRNA Gene Amplicon Sequence Data – Nicole A. Fiore – Microbiology Resource Announcements
Methods
A Customized At-Home Stool Collection Protocol for Use in Microbiome Studies Conducted in Cancer Patient Populations – Stephanie R. Hogue – Microbial Ecology
Effect of storage and DNA extraction method on 16S rRNA-profiled fecal microbiota in Japanese adults – Yuki Kawada – Journal of Clinical Biochemistry and Nutrition
Bioinformatics
𝑊∗𝑑Wd∗-test: robust distance-based multivariate analysis of variance – Bashir Hamidi – Microbiome
Identification and reconstruction of novel antibiotic resistance genes from metagenomes – Fanny Berglund – Microbiome
April 04, 2019
In today’s microbiome digest, multiple research and review articles focusing on the role of host-associated microbiomes in various diseases including Type 2 Diabetes, Obesity, Urolithiasis, Bronchial Asthma, Cancer Immunotherapy, Hepatic Glycan Storage, Uveitis, Schizophrenia and the Development of the immune system. Enjoy reading.
General microbiome
Harnessing microbiota interactions to produce bioactive metabolites: communication signals and receptor proteins – Juan F Martin – Current Opinion in Pharmacology
Review: Prevention of allergy with diverse and healthy microbiota:an update – Diana Chernikova – Current Opinion in Pediatrics
State of the Art: Pharmacomicrobiomics: The holy grail to variability in drug response? – Anukriti Sharma – Clinical Pharmacology and Therapeutics
Gut microbiome
The alteration of gut microbiota in newly diagnosed type 2 diabetic patients – Pei-Chi Chen – Nutrition
Defining Dysbiosis in Patients with Urolithiasis – Anna Zampini – Scientific Reports
Review: Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity – Veronica Lazar – Frontiers in Nutrition
Metabolic activity of intestinal microflora in patients with bronchial asthma – Vladimir Irashkin – Clinics and Practice
Pharmacologic and Nonpharmacologic Therapies for the Gut Microbiota in Type 2 Diabetes – Robert Caesar – Canadian Journal of Diabetes
Review: Modulation of cancer immunotherapy efficacy by gut microbiota – Huo S – Discovery Medicine
Hepatic glycogen storage diseases are associated to microbial dysbiosis – Karina Colonetti – PLOS ONE
Implicating Dysbiosis of the Gut Fungal Microbiome in Uveitis, an Inflammatory Disease of the Eye – Rajagopalaboopathi Jayasudha – Immunology and Microbiology
Cohort profile: The LoewenKIDS Study – life-course perspective on infections, the microbiome and the development of the immune system in early childhood – Cornelia Gottschick – International Journal of Epidemiology
Case Report: Decreased Clostridium Abundance after Electroconvulsive Therapy in the Gut Microbiota of a Patient with Schizophrenia – Misako Kanayama – Case Reports in Psychiatry
Reproductive tract microbiome
Effects of oral probiotic supplements on vaginal microbiota during pregnancy: a randomised, double‐blind, placebo‐controlled trial with microbiome analysis – S Husain – BJOG
Skin microbiome
Review: The Cutaneous Microbiome: Implications for Dermatology Practice – Megan A. Sander
Animal microbiome / Animal experiments
Intestinal microbiota of broilers submitted to feeding restriction and its relationship to hepatic metabolism and fat mass: Fast-growing strain – Raquel Lunedo – Journal of Animal Physiology and Animal Nutrition
Phytate degradation, intestinal microbiota, microbial metabolites and immune values are changed in growing pigs fed diets with varying calcium–phosphorus concentration and fermentable substrates – Charlotte Maria Elisabeth Heyer – Journal of Animal Physiology and Animal Nutrition
Review: The importance of scale in comparative microbiome research: New insights from the gut and glands of captive and wild lemurs – Lydia K. Greene – American Journal of Primatology
A dietary isothiocyanate-enriched moringa (Moringa oleifera) seed extract improves glucose tolerance in a high-fat-diet mouse model and modulates the gut microbiome – Asha Jaja-Chimedza – Journal of Functional Foods
Musca domestica Cecropin (Mdc) Alleviates Salmonella typhimurium-Induced Colonic Mucosal Barrier Impairment: Associating With Inflammatory and Oxidative Stress Response, Tight Junction as Well as Intestinal Flora – Lun Zhang – Frontiers in Microbiology
The Effects of Unfermented and Fermented Cow and Sheep Milk on the Gut Microbiota – Elizabeth A. Rettedal – Frontiers in Microbiology
Water and extremophile microbiome
Microbiota of the Hickey Run Tributary of the Anacostia River – Robin Cagle – Microbiology Resource Announcements
Bioinformatics
Review: Statistical Analysis of Metagenomics Data – M. Luz Calle – Genomics and Informatics
March 28, 2019
Today’s digest covers interesting reviews on different aspects of host-microbiome interactions, research articles about how structural variation in gut microbiome affects host health, characterization of a simplified microbial community to study microbial interaction with diet in mice and viral diversity in soil metagenomes. Happy reading!
General microbiome
Review – Microbiome: Our opponents or allies in healthcare and medicine – Horst A von Recum – Experimental Biology and Medicine
Review – To B12 or not to B12: Five questions on the role of cobalamin in host-microbial interactions – Carol A. Rowley – Plos Pathogens
Review – Can oral bacteria affect the microbiome of the gut? – Ingar Olsen – Journal of Oral Microbiology
Review – Inter-kingdom signaling between gut microbiota and their host – Qing Li – Cellular and Molecular Life Sciences
Human gut microbiome
Structural variation in the gut microbiome associates with host health – David Zeevi – Nature
Effects of fermentable high fiber diet supplementation on gut derived and conventional nitrogenous product in patients on maintenance hemodialysis: a randomized controlled trial – Hamid Tayebi Khosroshahi – Nutrition and Metabolism
Identification of the microbial diversity after fecal microbiota transplantation therapy for chronic intractable constipation using 16s rRNA amplicon sequencing – Tadashi Ohara – PlosOne
The microbiome and tuberculosis: state of the art, potential applications, and defining the clinical research agenda – Charissa C Naidoo – Lancet Respiratory medicine
Fecal Microbiota Transplantation: An Update on Clinical Practice – Kyeong Ok Kim – Clinical Endoscopy
Aryl hydrocarbon receptor and uremic toxins from gut microbiota in chronic kidney disease patients: Is there a relationship? – Jessyca Brito – Biochemistry
Animal experiments
Bacterial AB5 toxins inhibit the growth of gut bacteria by targeting ganglioside-like glycoconjugates – Robert T. Patry – Nature Communications
Simplified Intestinal Microbiota to Study Microbe-Diet-Host Interactions in a Mouse Model – Petia Kovatcheva-Datchary – Cell Reports
Pomegranate ellagitannin-gut microbial-derived metabolites, urolithins, inhibit neuroinflammation in vitro – Nicholas A. DaSilva – Nutritional Neuroscience
Endogenous Enterobacteriaceae underlie variation in susceptibility to Salmonella infection – Eric M. Velazquez – Nature Microbiology
Plant, root, soil microbiome
Thickness determines microbial community structure and function in nitrifying biofilms via deterministic assembly – Carolina Suarez – Scientific Reports
Untapped viral diversity in global soil metagenomes – Emily B. Graham – BioRxiv
Water microbiome and extremophiles
Fate of antibiotic resistance genes in two Arctic tundra wetlands impacted by municipal wastewater – Jennifer L. Hayward – Science of the Total Environment
Probiotics
Importance of probiotics in the prevention and treatment of colorectal cancer – Majid Eslami – Journal of Cellular Physiology
Non-microbiology picks
Artificial womb technology breaks its four minute mile – Science Daily
